
Abstract
Aims/hypothesis
A decrease in plasma adiponectin levels has been shown to contribute to the development of diabetes. However, it remains uncertain whether adiponectin plays a role in the regulation of insulin secretion. In this study, we investigated whether adiponectin may be involved in the regulation of insulin secretion in vivo and in vitro.
Methods
The effect of adiponectin on insulin secretion was measured in vitro and in vivo, along with the effects of adiponectin on ATP generation, membrane potentials, Ca2+ currents, cytosolic calcium concentration and state of 5′-AMP-activated protein kinase (AMPK). In addition, insulin granule transport was measured by membrane capacitance and total internal reflection fluorescence (TIRF) analysis.
Results
Adiponectin significantly stimulated insulin secretion from pancreatic islets to approximately 2.3-fold the baseline value in the presence of a glucose concentration of 5.6 mmol/l. Although adiponectin had no effect on ATP generation, membrane potentials, Ca2+ currents, cytosolic calcium concentrations or activation status of AMPK, it caused a significant increase of membrane capacitance to approximately 2.3-fold the baseline value. TIRF analysis revealed that adiponectin induced a significant increase in the number of fusion events in mouse pancreatic beta cells under 5.6 mmol/l glucose loading, without affecting the status of previously docked granules. Moreover, intravenous injection of adiponectin significantly increased insulin secretion to approximately 1.6-fold of baseline in C57BL/6 mice.
Conclusions/interpretation
The above results indicate that adiponectin induces insulin secretion in vitro and in vivo.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The adipocyte-derived hormone adiponectin (also known as Acrp30, GBP28 or AdipoQ) [1–4] has been shown to play important roles in the regulation of glucose and lipid metabolism. Plasma adiponectin levels are reduced in obese and insulin-resistant humans and in animal models [2, 5, 6]. Adiponectin improves insulin sensitivity in muscle and liver by enhancing fatty acid oxidation via activation of 5′-AMP-activated protein kinase (AMPK) [7, 8] and peroxisome proliferator-activated receptor α [6, 9].
We previously demonstrated that adiponectin-deficient (Adipo −/−) mice exhibit insulin resistance [10], but that after glucose loading the plasma insulin levels tended to be lower in Adipo −/− mice than in wild-type mice, suggesting that adiponectin may induce insulin secretion [10]. The adiponectin receptors ADIPOR1 and ADIPOR2 have recently been cloned [11] and identified in human and rat pancreatic beta cells [12]. Expression of the adiponectin receptors by INS-1 cells (a clonal rat beta cell line) has been found to increase following exposure to oleic acid, an unsaturated NEFA. Moreover, a previous study has suggested that adiponectin also exerts anti-apoptotic actions and that this protective function of adiponectin might serve to counteract autoimmune- and lipotoxicity-induced beta cell destruction [13]. However, it is still uncertain whether adiponectin plays a role in the regulation of insulin secretion. In this study we investigated whether adiponectin might be involved in the regulation of insulin secretion in vitro and in vivo.
Methods
Animals
C57BL/6 mice were obtained from CLEA Japan (Tokyo, Japan). Male C57BL/6 mice (10 to 16 weeks old) were housed under a 12 h light–dark cycle and given free access to food. The animal care and experimental procedures were approved by the Animal Care Committee of the University of Tokyo.
RNA preparation and real-time quantitative PCR
Total RNA was extracted from isolated islets using TRIzol reagent (Invitrogen, Carlsbad, CA, USA) according to the manufacturer’s instructions. cDNA synthesis was performed using the SuperScript Preamplification System (Invitrogen), followed by TaqMan quantitative PCR (Applied Biosystems, Foster City, CA, USA; 50°C for 2 min and 95°C for 10 min, followed by 40 cycles of 95°C for 15 s and 60°C for 1 min) with an ABI Prism 7000 PCR instrument (Applied Biosystems) to amplify samples of the Adipor1, Adipor2 and β-actin genes. The sequences of the primers and probes are described elsewhere [11].
Insulin secretion by islets
Islets were isolated from 10- to 16-week-old C57BL/6 mice as described elsewhere [14]. In brief, after clamping the common bile duct at a point close to the duodenal outlet, 2.5 ml KRB (129 mmol/l NaCl, 4.8 mmol/l KCl, 1.2 mmol/l MgSO4, 1.2 mmol/l KH2PO4, 2.5 mmol/l CaCl2, 5 mmol/l NaHCO3 and 10 mmol/l HEPES; pH 7.4) containing 4 mg/ml collagenase (Sigma, St Louis, MO, USA) was injected into the duct. Insulin release by the pancreatic islets was measured by static incubation with KRB containing 0.2% (wt/vol.) bovine serum albumin [15]. In the static incubation, batches of ten freshly isolated islets were preincubated at 37°C for 30 min in KRB containing 2.8 mmol/l glucose. The preincubation solutions were replaced with KRB containing test agents and the batches of islets were incubated at 37°C for 60 min. The insulin released in the supernatant fractions was then measured by radioimmunoassay (Biotrak; GE Healthcare, Chalfont St Giles, UK).
Generation of recombinant adiponectin
Bacterially expressed murine full-length adiponectin was generated as described previously [7, 11]. The endotoxin content of the purified protein was determined using a Limulus Amebocyte Lysate Assay (Bio Whittaler, Walkersville, MD, USA). Mammalian adiponectin was purchased from Alexis Biochemicals (San Diego, CA, USA).
Glucose oxidation and fatty acid oxidation
Glucose oxidation in the islets was evaluated by measuring the 14CO2 generation from d-[6–14C]glucose [16]. Batches of ten freshly isolated islets were incubated at 37°C for 60 min in KRB containing 29.6 kBq d-[6–14C]glucose and 5.6 or 22.2 mmol/l glucose with and without 10 μg/ml adiponectin. For fatty acid oxidation, batches of ten freshly isolated islets were incubated at 37°C for 60 min in KRB containing 25.9 kBq [U-14C]palmitic acid, 1 mmol/l carnitine, and 5.6 or 22.2 mmol/l glucose with and without 10 μg/ml adiponectin. The 14CO2 produced was volatilised by adding HCl, captured with Solvable (Packard Instrument Company, Meriden, CT, USA) and measured by liquid scintillation counting.
ATP and cAMP content
The ATP and cAMP contents of the islets were determined as described previously [17]. Briefly, batches of ten islets were incubated at 37°C for 60 min in KRB containing 5.6 mmol/l glucose with and without 10 μg/ml adiponectin. Incubation was stopped by the addition of ice-cold HClO4 and the islets were homogenised by sonication. The lysates were neutralised by the addition of NaOH. The ATP content and cAMP content of the supernatant fraction were measured using a bioluminescent assay kit (Sigma) and an enzyme-linked immunoassay kit (GE Healthcare), respectively.
Electrical recordings
Electrophysiological experiments on the islets were performed using cells in situ in intact pancreatic islets. The islets were washed extensively in collagenase-free solution and then maintained in a short-term tissue culture (<24 h) in RPMI 1640 containing 10% (vol./vol.) fetal calf serum supplemented with 100 μg/ml streptomycin and 100 IU/ml penicillin.
To establish the whole-cell mode, the amphotericin B-perforated patch-clamp technique was used. The membrane potential and the current of superficial cells in the intact pancreatic islets were recorded using an EPC-9 patch-clamp amplifier (HEKA Electronics, Lambrecht/Pfalz, Germany). The beta cells were identified by the lack of inward Na+ currents in the presence of a depolarising pulse from −70 to 0 mV (5 ms in duration) [18]. The capacitance measurements were performed using software-based lock-in software (Pulse version 8.11; HEKA Electronics). The changes in cell capacitance were estimated by the Lindau–Neher technique [19, 20] by implementing the ‘Sine + DC’ feature of the lock-in module. The amplitude of the sine wave was 20 mV and the frequency was set at 1250 Hz. Patch pipettes (tip resistance 5–7 MΩ when filled with the pipette solution) were pulled from borosilicate tubing. The capacitance measurements were performed at 32–34°C. Capacitance was measured again 2 min 30 s after treatment with or without adiponectin. The standard extracellular medium consisted of 120 mmol/l NaCl, 20 mmol/l tetraethylammonium-Cl, 3.6 mmol/l KCl, 2 mmol/l NaHCO3, 0.5 mmol/l NaH2PO4, 0.5 mmol/l MgSO4, 5 mmol/l HEPES (at pH 7.4), 2.6 mmol/l CaCl2 and 5 mmol/l d-glucose. The pipette solution for the perforated patch was composed of 76 mmol/l Cs2SO4, 10 mmol/l NaCl, 10 mmol/l KCl, 1 mmol/l MgCl2 and 5 mmol/l HEPES (at pH 7.35).
The cytosolic Ca2+ concentration ([Ca2+]c) was measured using fura-2 by exciting its fluorescence in a dual-wavelength ratiometric mode at 340 and 380 nm. The emission wavelength was filtered at 500 nm. [Ca2+]c was expressed as the 340:380 nm ratio.
Western blot analysis
Mouse islets were homogenised in ice-cold buffer A (25 mmol/l Tris-HCl; pH 7.4, 10 mmol/l sodium orthovanadate, 10 mmol/l sodium pyrophosphate, 100 mmol/l sodium fluoride, 10 mmol/l EDTA, 10 mmol/l EGTA and 1 mmol/l phenylmethylsulfonyl fluoride). Samples were separated on polyacrylamide gels and transferred to a nitrocellulose membrane (Schleicher & Schuell, Dassel, Germany). Rabbit anti-phospho-AMPKα (Thr-172) antibody and rabbit AMPKα antibody were purchased from Cell Signaling (Beverly, MA, USA).
Total internal reflection fluorescence microscopy
We performed total internal reflection fluorescence (TIRF) microscopy using an inverted microscope (IX70; Olympus, Tokyo, Japan) with a high-aperture objective lens (Apo 100× OHR, Na 1.65; Olympus) as described previously [21, 22]. Mouse pancreatic islets were isolated by collagenase digestion. Isolated islets were dissociated into single cells by incubation in Ca2+-free KRB containing 1 mmol/l EGTA and the beta cells were cultured on high-refractive-index glass (Olympus) coated with fibronectin (Koken, Tokyo, Japan). To label the insulin-containing granules, pancreatic beta cells were infected with recombinant adenovirus Adex1CA insulin green fluorescent protein (GFP) as described previously [22]. The experiments were performed 2 days after infection. To observe GFP, we used a 488 nm laser line for excitation. The procedure used to monitor the motion of GFP-labelled insulin granules in pancreatic beta cells by TIRF microscopy has been described elsewhere [21]. Infected cells on a glass coverslip (Olympus) were mounted in an open chamber and incubated for 30 min at 37°C in KRB and 0.3% (wt/vol.) bovine serum albumin in the presence of 2.8 mmol/l glucose. The cells were then transferred to a thermostatically controlled stage (37°C) and stimulated with 10 μg/ml adiponectin in the presence of 5.6 mmol/l glucose in the chamber. Diodomethane sulphur immersion oil (Cargille Laboratories, Cedar Grove, NJ, USA) was used to establish contact between the objective lens and the coverslip. The space constant for the exponential decay of the evanescent field was approximately 43 nm.
Image acquisition and analysis
Images were acquired every 300 ms with a cooled charge-coupled-device camera (DV887DCSBV; Andor Technology, South Windsor, CT, USA) operated with Metamorph 6.2 software (Universal Imaging, Downingtown, PA, USA). Most analyses, including tracking (single projection of difference images) and area calculations, were performed with the Metamorph software. To analyse the data, fusion events were selected manually and the average fluorescence intensity of the individual granules in a 1 × 1 μm square placed over the granule centre was calculated. The number of fusion events was counted manually while looping approximately 15,000 time-lapsed frames. The sequences were exported as single TIRF files and further processed using Adobe Photoshop 6.0 software or converted into Quick Time movies.
Insulin secretion study in vivo
Four days before the insulin secretion study, a catheter consisting of a silicone part (Phicon Tube; Fuji Systems, Tokyo, Japan) and a polyethylene part (PE-50; BD Biosciences, Franklin Lakes, NJ, USA) was inserted into the right jugular vein of animals under general anaesthesia with sodium pentobarbital, to administer infusions. The studies were performed on the mice under conscious, unstressed conditions after 24 h fast. d-Glucose (0.2 g/kg) was injected intravenously through the catheter, either alone or with 0.6 mg/kg adiponectin. Blood samples were collected into a heparinised tube before and 10 and 20 min after the injection. After immediate centrifugation, the plasma was separated and stored at −20 °C until analysed.
Insulin levels were determined by radioimmunoassay. The insulin to glucose ratio was calculated using the formulas: (10 min insulin level − fasting insulin level)/(10 min blood glucose level − fasting blood glucose) or (20 min insulin level − fasting insulin level)/(20 min blood glucose level − fasting blood glucose level). The adiponectin levels were determined with a mouse adiponectin ELISA kit (Otsuka, Tokyo, Japan). All data were obtained from six independent experiments.
Statistical analysis
The statistical significance of differences between groups was determined using Student’s t test for unpaired comparisons, the Welch test, Dunnett’s t test or Steel’s z test. A p value of <0.05 was regarded as significant.
Results
Adiponectin stimulates insulin secretion from mouse pancreatic islets at low glucose concentration
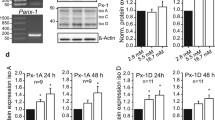
We first confirmed the expression of Adipor1 and Adipor2 in mouse pancreatic islets (Fig. 1a,b), obtaining results that were essentially consistent with those previously reported [12]. We next examined the effect of adiponectin on glucose-induced insulin secretion from isolated islets during a 60 min static incubation (Fig. 1c). Adiponectin significantly stimulated insulin secretion to 2.3-fold the baseline value at a 5.6 mmol/l glucose concentration (p = 0.012; Fig. 1c). However, adiponectin did not significantly increase insulin secretion in the presence of 11.1 or 22.2 mmol/l glucose. Since high concentrations of lipopolysaccharide (LPS) have been reported to stimulate insulin secretion from islets [23], we investigated the effect of LPS on insulin secretion from islets at 5.6 mmol/l glucose. When mouse pancreatic islets were treated with LPS at the same quantity as was present in recombinant adiponectin derived from Escherichia coli, no effect of LPS was found on insulin secretion (Fig. 1d). In addition, 10 μg/ml mammalian-derived adiponectin also stimulated insulin secretion from mouse pancreatic islets (p = 0.039; Fig. 1e). These results suggest that adiponectin acts directly on beta cells to increase insulin secretion.

Adiponectin stimulates insulin secretion from islets. TaqMan RT-PCR of Adipor1 (n = 3) (a) and Adipor2 (n = 3) (b) in mouse pancreatic islets. c Islets were incubated in KRB containing the indicated concentrations of glucose with or without adiponectin (n = 9–20). d Islets were incubated with glucose and with adiponectin or LPS (n = 3). e Islets were incubated with glucose with or without mammalian-derived adiponectin (n = 3). Values are means±SEM. *p < 0.05
Adiponectin stimulates insulin secretion without causing ATP generation, palmitic acid oxidation or cAMP generation in islets
To examine the effect of adiponectin on ATP generation, we measured glucose-induced changes in d-[6–14C]glucose oxidation and the ATP content of islets (Table 1). d-[6–14C]Glucose oxidation to 14CO2 in the control islets increased 5.5-fold when the extracellular glucose concentration was raised from 5.6 to 22.2 mmol/l. Adiponectin at a concentration of 10 μg/ml did not affect d-[6–14C]glucose oxidation in the presence of 5.6 mmol/l of glucose compared with control. In the presence of 22.2 mmol/l glucose, adiponectin also failed to significantly change d-[6–14C]glucose oxidation compared with control islets incubated at the same glucose concentration. Next, we measured the ATP content of islets to directly monitor the efficiency of mitochondrial ATP synthesis. The results showed that adiponectin at a concentration of 10 μg/ml did not affect ATP content in the presence of 5.6 mmol/l glucose (Table 1). The above findings indicate that adiponectin has no stimulatory effect on ATP generation from mitochondrial glucose metabolism. Increase in cAMP content potentiates glucose-stimulated insulin secretion through activation of cAMP-dependent protein kinase [24]. Adiponectin also failed to significantly change the cAMP content as compared with control islets (Table 1). Although adiponectin has been reported to reduce elevated fatty acid levels in muscle by oxidising fatty acids [7–9], no significant change in the oxidation of palmitic acid was observed in pancreas islets treated with adiponectin as compared with control at 5.6 or 22.2 mmol/l of glucose (Table 1).
Adiponectin stimulates insulin secretion without causing membrane depolarisation, closure of \({\text{K}}_{{\text{ATP}}}^ + - {\text{channels}}\) or Ca2+ entry into the cytosol
We measured the membrane potentials of pancreatic beta cells to determine whether adiponectin caused membrane depolarisation. In the presence of 5.6 mmol/l glucose, adiponectin at a concentration of 10 μg/ml had no effect on the membrane potentials (Fig. 2a), but membrane of pancreatic beta cells depolarised when the extracellular glucose level was raised from 5.6 to 22.2 mmol/l (Fig. 2a). Figure 2b shows the current–voltage relationship for Ca2+ currents. When the membrane potential was held at −70 mV, Ca2+ currents elicited by 200 ms depolarising voltage-clamp pulses from −40 to 40 mV were applied in the presence of 5.6 mmol/l glucose. Adiponectin at a concentration of 10 μg/ml did not change the current–voltage curve via Ca2+ influx into the cytosol (Fig. 2b). To investigate the effect of adiponectin on the increase in the [Ca2+]c, we monitored the fluorescence of fura-2/acetoxymethyl ester excited at 340 and 380 nm in perifused islets (Fig. 2c). The 340:380 nm fluorescence ratio increased in islets when the glucose concentration was raised from 5.6 to 22.2 mmol/l, but adiponectin had no effect on the [Ca2+]c (Fig. 2c). To clarify whether adiponectin stimulates insulin secretion without causing Ca2+ influx, we next investigated the effects of adiponectin on mouse pancreatic islets in the presence of the L-type Ca2+-channel blocker, nitrendipine. Nitrendipine at a concentration of 5 μmol/l is known to inhibit Ca2+ influx [25]. Nevertheless, insulin secretion was markedly diminished at 22.2 mmol/l glucose in the presence of nitrendipine, whereas adiponectin significantly stimulated insulin secretion at 5.6 mmol/l glucose (p = 0.043; Fig. 2d). Interestingly, adiponectin also significantly stimulated insulin secretion at 22.2 mmol/l glucose concentration in the presence of nitrendipine (p = 0.047) (Fig. 2d). These findings suggest that adiponectin is able to stimulate insulin secretion when the influx of Ca2+ through voltage-dependent Ca2+ channels is blocked.

Adiponectin stimulates insulin secretion without causing membrane depolarisation, closure of \({\text{K}}_{{\text{ATP}}}^ + - {\text{channels}}\)or Ca2+ entry into the cytosol. a Membrane potentials in response to the addition of adiponectin and increasing concentrations of glucose (5.6–22.2 mmol/l) were recorded under the conventional whole-cell mode. Results are representative of three independent experiments. b The current–voltage relationships of beta cells treated with (black circles, n = 3) or without (white circles, n = 3) adiponectin (10 μg/ml). c Cytosolic calcium concentrations in response to the addition of adiponectin and increasing concentrations of glucose were measured using fura-2. Mean values are shown (n = 5). d Islets were incubated in KRB containing the indicated concentrations of glucose and 5 μmol/l nitrendipine with or without 10 μg/ml adiponectin (n = 18–23). Values are means ± SEM. *p < 0.05
AMPK is not involved in adiponectin-stimulated insulin secretion
Adiponectin has been previously reported to improve insulin sensitivity in muscle and liver by enhancing fatty acid oxidation via the activation of AMPK [7, 8], which has been found to increase glucose transport by stimulating the translocation of GLUT4 [26, 27]. To elucidate whether AMPK might be involved in adiponectin-stimulated insulin secretion, we examined the effect of adiponectin on phosphorylation of AMPK in mouse pancreatic islets. However, adiponectin did not affect the phosphorylation of AMPK at 5.6 mmol/l glucose (Fig. 3a). The AMPK activator 5-amino-imidazole-4-carboxamide riboside (AICAR), on the other hand, significantly increased the phosphorylation of AMPK at 5.6 mmol/l glucose (p = 0.00083; Fig. 3b). However, AICAR significantly decreased insulin secretion from mouse pancreatic islets (p = 0.024; Fig. 3c) [28]. These results suggest that AMPK is not involved in adiponectin-stimulated insulin secretion at low glucose concentrations.

Adiponectin stimulates insulin secretion without causing activation of AMPK. a, b Western blots of phosphorylated AMPK and AMPKα in mouse pancreatic islets (n = 3, each group). Lysates from 30 islets from each group were subjected to western blot analysis. c Isolated islets were incubated with AICAR at 5.6 mmol/l glucose. Values are means±SEM. *p < 0.05, ***p < 0.001
Adiponectin stimulates insulin release via induction of increased fusion events at a low glucose concentration
We then examined the capacitance of mouse pancreatic beta cells. The technique used is characterised by high time resolution and thus can detect dynamic changes in the cell surface area that reflect exocytosis. Figure 4a shows the capacitance of mouse pancreatic beta cells. Interestingly, adiponectin at a concentration of 10 μg/ml significantly increased the capacitance of mouse pancreatic beta cells to 2.3-fold above baseline (p = 0.0091; Fig. 4a), indicating that adiponectin may stimulate insulin granule exocytosis in the absence of any changes in membrane depolarisation, Ca2+ currents or [Ca2+]c.

Adiponectin stimulates insulin release via induction of increased fusion events. a The membrane capacitance of mouse beta cells exposed to 5.6 mmol/l glucose alone (lower curve) and 5.6 mmol/l glucose + 10 μg/ml adiponectin (upper curve) cumulatively increased upon repetitive application of depolarising pulses and then plateaued (glucose alone). The traces shown are representative of three experiments. b, c TIRF images were acquired every 300 ms after the addition of adiponectin. The fusion events per 200 μm2 were counted manually. Histograms of the number of fusion events in 5.6 mmol/l glucose in the absence (b) (n = 45) and presence (c) (n = 52) of adiponectin (10 μg/ml). Black bars, previously docked granules; white bars, newly docked granules. Values are mean±SEM. *p < 0.05, **p < 0.01, ***p < 0.005
To elucidate the mechanism by which adiponectin increased the exocytosis of insulin-containing granules, we used TIRF microscopy to monitor the real-time docking and fusion process of single insulin granules labelled with a GFP-tagged insulin near the plasma membrane in mouse pancreatic beta cells. Interestingly, adiponectin significantly increased the number of fusion events by newcomers (40.5 ± 4.1 per 200 μm2 in 0–30 min control beta cells vs 72.3 ± 6.5 per 200 μm2 in 0–30 min adiponectin-treated beta cells, p < 0.0005, n = 45 for control and n = 52 for adiponectin-treated cells; Fig. 4b,c and Electronic supplementary material [ESM], video clips), suggesting that adiponectin may accelerate transport of insulin granules from a cytoplasmic pool to the plasma membrane.
Adiponectin stimulates insulin secretion in vivo
To elucidate the physiological roles of adiponectin in insulin secretion in vivo, we studied the effect of adiponectin on insulin secretion in C57BL/6 mice after intravenous transcatheter injection of adiponectin and a low concentration of glucose. We tried to adjust the blood glucose level to approximately 5.6 mmol/l. Figure 5a shows the plasma adiponectin level at 10 and 20 min after the intravenous injection of glucose and saline or adiponectin. The plasma adiponectin levels increased significantly to 1.8 fold of baseline following the injection of adiponectin, while the increased blood glucose levels at 10 and 20 min after the injection were not significantly different between the two groups (Fig. 5b). The blood glucose level and insulin level at the starting point were 3.7 ± 0.4 mmol/l and 36.2 ± 5.0 pmol/l, respectively. No significant differences in the increased plasma insulin levels at 10 min after administration were observed between the adiponectin and the saline groups (Fig. 5c). However, the increase in plasma insulin levels at 20 min after the intravenous injection of glucose was approximately 1.6-fold greater in the adiponectin group (n = 19) than in the saline group (n = 21; p = 0.048; Fig. 5c). We evaluated the changes in ratio of the increment of plasma insulin level to that of blood glucose level. At 20 min after glucose loading the ratio in the adiponectin group was approximately sevenfold higher than that in the saline group (p = 0.0088) (Fig. 5d). These results suggest that adiponectin stimulates insulin secretion in vivo.

Adiponectin stimulates insulin secretion in vivo. a Plasma adiponectin level after intravenous injection of glucose and saline (white bars, n = 8–21) or adiponectin (black bars, n = 8–19). b Blood glucose and (c) plasma insulin levels after the intravenous injections. d Ratio of the increment in plasma insulin level to that of blood glucose level after the intravenous injections. Values are means±SEM. *p < 0.05, **p < 0.01, ***p < 0.005
Discussion
In this study we have shown that adiponectin stimulates insulin secretion in vitro and in vivo. In addition, our results also indicate that adiponectin stimulates insulin secretion without causing ATP generation, closure of \({\text{K}}_{{\text{ATP}}}^ + - {\text{channels}}\), Ca2+ entry into the cytosol or activation of AMPK.
Adiponectin has previously been reported to increase fatty acid oxidation via activation of AMPK [7, 8], which has been found to increase glucose transport by stimulating the translocation of GLUT4 to the sarcolemma in heart [26] and skeletal muscle [27]. However, adiponectin did not affect the phosphorylation of AMPK in pancreatic islets at 5.6 mmol/l glucose. Since AMPK in pancreatic beta cells is activated by low glucose concentrations [28], adiponectin may be unable to further activate AMPK at this low glucose concentration. By contrast, although the AMPK activator AICAR activated AMPK at this low glucose concentration, AICAR also decreased insulin secretion under these conditions. These results indicate that AMPK is not involved in adiponectin-stimulated insulin secretion at a low glucose concentration.
This study has shown that adiponectin stimulated insulin secretion without causing Ca2+ entry into the cytosol. The following cascade is generally accepted to be involved in glucose-induced insulin secretion. When glucose is metabolised in the cytosol and mitochondria, ATP is generated and promotes the closure of ATP-sensitive potassium channels, thereby depolarising the plasma membrane potential. Depolarisation of the plasma membrane leads to activation of voltage-dependent Ca2+ channels, Ca2+ entry into the cytosol and a rise in [Ca2+]c, which is thought to finally trigger the exocytosis of insulin-containing granules [29–31]. However, a Ca2+-independent pathway has also been proposed. Thus, Komatsu et al. have suggested that both Ca2+-dependent and Ca2+-independent augmentation occurs via a pathway dependent on glucose metabolism [32, 33]. In this context, we confirmed that adiponectin is able to stimulate insulin secretion when the influx of Ca2+ through voltage-dependent Ca2+ channels is blocked by nitrendipine; in other words, adiponectin stimulates insulin secretion without requiring influx of Ca2+ through voltage-dependent Ca2+ channels. It may do this through these augmentation pathways triggered by glucose metabolism [33], namely by replacing part of the Ca2+-dependent/ Ca2+-independent augmentation on newly docked granules. In this regard, it should be noted that adiponectin was able to stimulate insulin secretion at 22.2 mmol/l glucose when the influx of Ca2+ was blocked by nitrendipine (Fig. 2d). The observation that this stimulation of insulin secretion by adiponectin is relatively small as compared with that of glucose itself under physiological conditions (Fig. 1c) may reflect the relatively small role of the Ca2+-independent augmentation pathway, as suggested by Sato et al. [34].
As another possible mechanism, adiponectin may affect remodelling of the cytoskeleton. The cytoskeleton plays a critical role in glucose-stimulated intracellular trafficking and the exocytosis of insulin-containing granules at the plasma membrane of beta cells. In fact, a previous report showed direct evidence of a dynamic interaction between target membrane soluble N-ethylmaleimide-sensitive factor attachment protein receptor protein and F-actin during glucose-stimulated insulin secretion in MIN6 cells [35]. It was recently reported that calpain plays a role in facilitating the actin reorganisation required for glucose-stimulated insulin secretion [36]. Another study has suggested that kinesin I plays an important role in the microfilament-dependent movements of insulin containing granules [37]. Adiponectin may cause direct activation of calpain or kinesin I, to influence remodelling of the cytoskeleton.
Conflicting results on the effects of adiponectin on insulin secretory function in vitro have been reported so far. A previous study showed that while adiponectin decreased glucose/forskolin-induced insulin secretion, it reversed NEFA-induced inhibition of insulin secretion [13]. Another group reported that adiponectin had no effect on either basal or stimulated insulin secretion from human islets [38]. Winzell et al. [39] reported that adiponectin decreases insulin secretion from the pancreatic islets of insulin-resistant mice at a 2.8 mmol/l glucose, but stimulates insulin secretion at 16.7 mmol/l glucose. However, there have been no reports on the effects of adiponectin on insulin secretion in vivo. This study is the first study to demonstrate that adiponectin stimulates insulin secretion not only in vitro, but also in vivo.
In conclusion, we have described a novel effect of adiponectin, namely stimulation of insulin secretion, in addition to its known insulin-sensitising and anti-atherogenic effects. Consequently, adiponectin receptor agonists may be useful in the activation of adiponectin signalling as a new therapeutic strategy for treating diabetes and atherogenic diseases.
Abbreviations
- AICAR:
-
5-amino-imidazole-4-carboxamide riboside
- AMPK:
-
5′-AMP-activated protein kinase
- [Ca2+]c :
-
cytosolic Ca2+ concentration
- GFP:
-
green fluorescent protein
- LPS:
-
lipopolysaccharide
- TIRF:
-
total internal reflection fluorescence
References
Scherer PE, Williams S, Fogliano M, Baldini G, Lodish HF (1995) A novel serum protein similar to C1q, produced exclusively in adipocytes. J Biol Chem 270:26746–26749
Hu E, Liang P, Spiegelman BM (1996) AdipoQ is a novel adipose-specific gene dysregulated in obesity. J Biol Chem 271:10697–10703
Maeda K, Okubo K, Shimomura I, Funahashi T, Matsuzawa Y, Matsubara K (1996) cDNA cloning and expression of a novel adipose specific collagen-like factor, apM1 (AdiPose Most abundant Gene transcript 1). Biochem Biophys Res Commun 221:286–296
Nakano Y, Tobe T, Choi-Miura NH, Mazda T, Tomita M (1996) Isolation and characterization of GBP28, a novel gelatin-binding protein purified from human plasma. J Biochem (Tokyo) 120:803–812
Arita Y, Kihara S, Ouchi N et al (1999) Paradoxical decrease of an adipose-specific protein, adiponectin, in obesity. Biochem Biophys Res Commun 257:79–83
Yamauchi T, Kamon J, Waki H et al (2001) The fat-derived hormone adiponectin reverses insulin resistance associated with both lipoatrophy and obesity. Nat Med 7:941–946
Yamauchi T, Kamon J, Minokoshi Y et al (2002) Adiponectin stimulates glucose utilization and fatty-acid oxidation by activating AMP-activated protein kinase. Nat Med 8:1288–1295
Tomas E, Tsao TS, Saha AK et al (2002) Enhanced muscle fat oxidation and glucose transport by ACRP30 globular domain: acetyl-CoA carboxylase inhibition and AMP-activated protein kinase activation. Proc Natl Acad Sci U S A 99:16309–16313
Fruebis J, Tsao TS, Javorschi S et al (2001) Proteolytic cleavage product of 30-kDa adipocyte complement-related protein increases fatty acid oxidation in muscle and causes weight loss in mice. Proc Natl Acad Sci U S A 98:2005–2010
Kubota N, Terauchi Y, Yamauchi T et al (2002) Disruption of adiponectin causes insulin resistance and neointimal formation. J Biol Chem 277:25863–25866
Yamauchi T, Kamon J, Ito Y et al (2003) Cloning of adiponectin receptors that mediate antidiabetic metabolic effects. Nature 423:762–769
Kharroubi I, Rasschaert J, Eizirik DL, Cnop M (2003) Expression of adiponectin receptors in pancreatic beta cells. Biochem Biophys Res Commun 312:1118–1122
Rakatzi I, Mueller H, Ritzeler O, Tennagels N, Eckel J (2004) Adiponectin counteracts cytokine- and fatty acid-induced apoptosis in the pancreatic beta-cell line INS-1. Diabetologia 47:249–258
Lacy PE, Malaisse WJ (1973) Microtubules and beta cell secretion. Recent Prog Horm Res 29:199–228
Eto K, Tsubamoto Y, Terauchi Y et al (1999) Role of NADH shuttle system in glucose-induced activation of mitochondrial metabolism and insulin secretion. Science 283:981–985
Schuit F, De Vos A, Farfari S et al (1997) Metabolic fate of glucose in purified islet cells. Glucose-regulated anaplerosis in beta cells. J Biol Chem 272:18572–18579
Malaisse WJ, Sener A (1987) Glucose-induced changes in cytosolic ATP content in pancreatic islets. Biochim Biophys Acta 927:190–195
Gopel S, Zhang Q, Eliasson L et al (2004) Capacitance measurements of exocytosis in mouse pancreatic alpha-, beta-, and delta-cells within intact islets of Langerhans. J Physiol 556:711–726
Gillis KD (1995) Single-channel recording. Plenum, New York, pp 155–197
Kanno T, Ma X, Barg S et al (2004) Large dense-core vesicle exocytosis in pancreatic beta-cells monitored by capacitance measurements. Methods 33:302–311
Ohara-Imaizumi M, Nakamichi Y, Tanaka T, Ishida H, Nagamatsu S (2002) Imaging exocytosis of single insulin secretory granules with evanescent wave microscopy: distinct behavior of granule motion in biphasic insulin release. J Biol Chem 277:3805–3808
Ohara-Imaizumi M, Nishiwaki C, Kikuta T, Nagai S, Nakamichi Y, Nagamatsu S (2004) TIRF imaging of docking and fusion of single insulin granule motion in primary rat pancreatic beta-cells: different behaviour of granule motion between normal and Goto-Kakizaki diabetic rat beta-cells. Biochem J 381:13–18
Vives-Pi M, Somoza N, Fernandez-Alvanrez J et al (2003) Evidence of expression of endotoxin receptors CD14, toll-like receptors TLR4 and TLR2 and associated molecule MD-2 and of sensitivity to endotoxin (LPS) in islet beta cells. Clin Exp Immunol 133:208–218
Eddlestone GT, Oldham SB, Lipson LG, Premdas FH, Beigelman PM (1985) Electrical activity, cAMP concentration, and insulin release in mouse islets of Langerhans. Am J Physiol 248:C145–C153
Ikeda Y, Iguchi H, Nakata M et al (2005) Identification of N-arachidonylglycine, U18666A, and 4-andorostene-3, 17-dione as novel insulin secretagogues. Biochem Biophys Res Commun 333:778–786
Russell RR 3rd, Bergeron R, Shulman GI, Young LH (1999) Translocation of myocardial GLUT4 and increased glucose uptake through activation of AMPK by AICAR. Am J Physiol 277:H643–H649
Kurth-Kraczek EJ, Hirshman MF, Goodyear LJ, Winder WW (1999) 5′ AMP-activated protein kinase activation causes GLUT4 translocation in skeletal muscle. Diabetes 48:1667–1671
da Silva Xavier G, Leclerc I, Varadi A, Tsuboi T, Moule SK, Rutter GA (2003) Role for AMP-activated protein kinase in glucose-stimulated insulin secretion and preproinsulin gene expression. Biochem J 371:761–774
Ashcroft FM, Proks P, Smith PA, Ammala C, Bokvist K, Rorsman P (1994) Stimulus-secretion coupling in pancreatic beta cells. J Cell Biochem 55:54–65
Dukes ID, Philipson LH (1996) K+ channels: generating excitement in pancreatic beta-cells. Diabetes 45:845–853
Wollheim CB, Lang J, Regazzi R (1996) The exocytotic process of insulin secretion and its regulation by Ca2+ and G-proteins. Diabetes Rev 4:276–297
Komatsu M, Schermerhorn T, Aizawa T, Sharp GW (1995) Glucose stimulation of insulin release in the absence of extracellular Ca2+ and in the absence of any increase in intracellular Ca2+ in rat pancreatic islets. Proc Natl Acad Sci U S A. 92:10728–10732
Komatsu M, Noda M, Sharp GW (1998) Nutrient augmentation of Ca2+-dependent and Ca2+-independent pathways in stimulus-coupling to insulin secretion can be distinguished by their guanosine triphosphate requirements: studies on rat pancreatic islets. Endocrinology 139:1172–1183
Sato Y, Nenquin M, Henquin JC (1998) Relative contribution of Ca2+-dependent and Ca2+-independent mechanisms to the regulation of insulin secretion by glucose. FEBS Lett 421:115–119
Thurmond DC, Gonelle-Gispert C, Furukawa M, Halban PA, Pessin JE (2003) Glucose-stimulated insulin secretion is coupled to the interaction of actin with the t-SNARE (target membrane soluble N-ethymaleimide-sensitive factor attachment protein receptor protein) complex. Mol Endocinol 17:732–742
Turner MD, Fulcher FK, Jones CV et al (2007) Calpain facilitates actin reorganization during glucose-stimulated insulin secretion. Biochem Biophys Res Commun 352:650–655
Varadi A, Tsuboi T, Johnson-Cadwell LI, Allan VJ, Rutter GA (2003) Kinesin I and cytoplasmic dynein orchestrate glucose-stimulated insulin-containing vesicle movements in clonal MIN6 beta-cells. Biochem Biophys Res Commun 311:272–282
Staiger K, Stefan N, Staiger H et al (2005) Adiponectin is functionally active in human islets but does not affect insulin secretory function or beta-cell lipoapoptosis. J Clin Endocrinol Metab 90:6707–6713
Winzell MS, Nogueiras R, Dieguez C, Ahren B (2004) Dual action of adiponectin on insulin secretion in insulin-resistant mice. Biochem Biophys Res Commun 321:154–160
Acknowledgements
We thank A. Ohyama, C. Kokaki-Kaizuka, N. Kowatari-Otsuka, K. Takasawa and H. Chiyonobu for their excellent technical assistance and mouse husbandry.
Duality of interest
The authors declare that there is no duality of interest associated with this manuscript.
Author information
Authors and Affiliations
Corresponding author
Additional information
M. Okamoto and M. Ohara-Imaizumi contributed equally to this study.
Electronic supplementary materials
Below is the linked to the electronic supplementary materials.
Rights and permissions
About this article
Cite this article
Okamoto, M., Ohara-Imaizumi, M., Kubota, N. et al. Adiponectin induces insulin secretion in vitro and in vivo at a low glucose concentration. Diabetologia 51, 827–835 (2008). https://doi.org/10.1007/s00125-008-0944-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00125-008-0944-9